
Centrale Physique 1 PC 2008
| Thème de l'épreuve | Biophysique de la bactérie E. Coli. |
| Principaux outils utilisés | mécanique des fluides, diffusion, électrostatique |
| Mots clefs | bactérie, flagelle, potentiel de membrane, force de traînée, moteur pas à pas, biologie |
Corrigé
: 👈 gratuite pour tous les corrigés si tu crées un compte👈 l'accès aux indications de tous les corrigés ne coûte que 5 € ⬅ clique ici
👈 gratuite pour tous les corrigés si tu crées un compte
- - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
Énoncé complet
(télécharger le PDF)











Rapport du jury
(télécharger le PDF)


Énoncé obtenu par reconnaissance optique des caractères
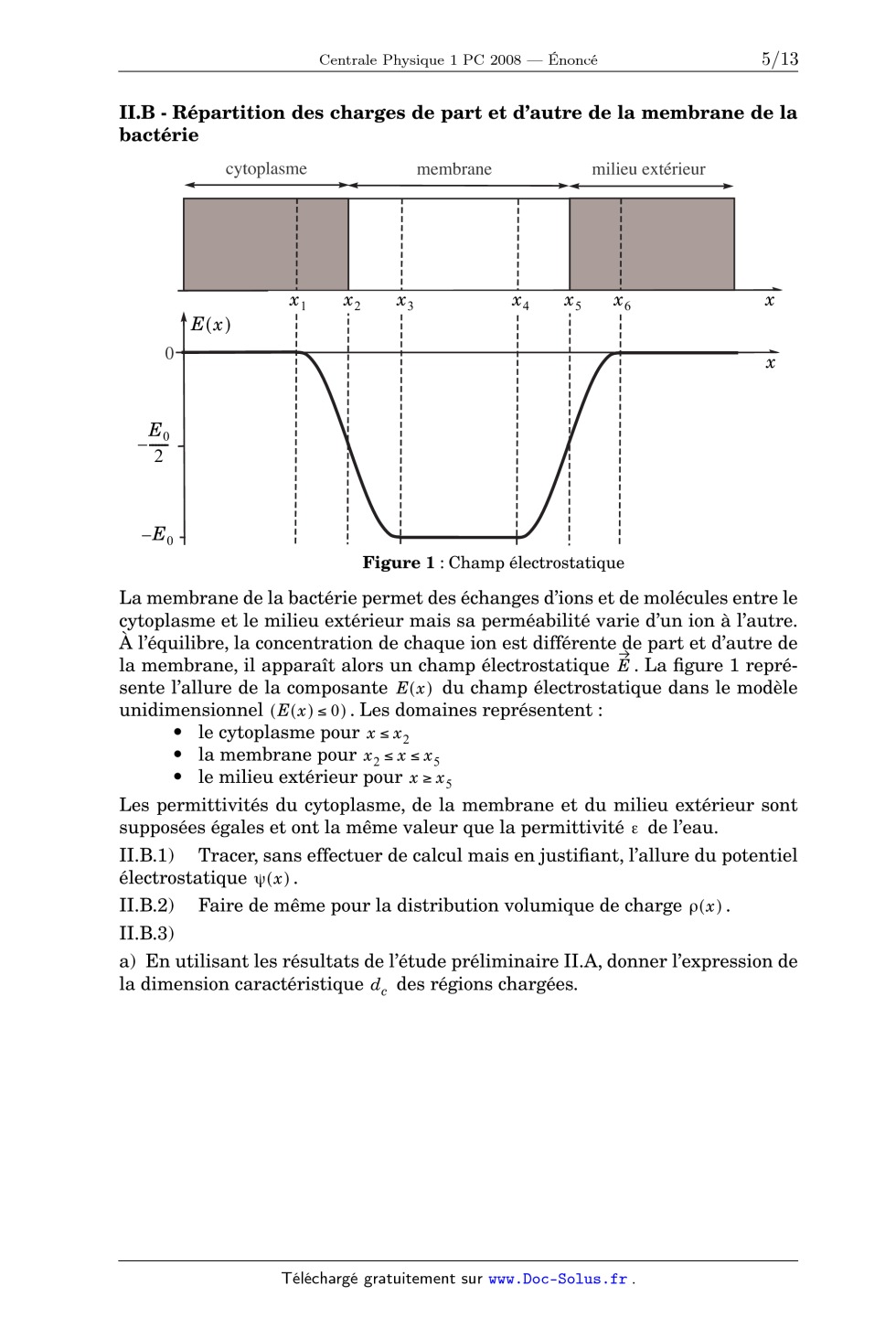
Concours Centrale - Supélec 2008 Épreuve : PHYSIQUE I Filière PC PHYSIQUE I Filière PC PHYSIQUE I Calculatrices autorisées. Biophysique de la bactérie Escherichia coli L'objet de ce problème est d'appliquer les lois de la physique à la bactérie Escherichia coli, que l'on nommera par la suite E. coli. Aucune connaissance de biolo6 9 gie n'est nécessaire. On rappelle que 1µm = 1 10 m et 1nm = 1 10 m . On rappelle également les valeurs : de la constante de Boltzmann : de la constante d'Avogadro : k B = 1, 38 10 NA 23 = 6, 02 10 de la permittivité du vide : 0 = 8, 85 10 de la charge élémentaire : e = 1, 6 10 23 12 19 JK mol Fm 1 1 1 C Les quatre parties de ce problème sont indépendantes. Partie I - Taille critique d'une bactérie aérobie On étudie les conditions de survie d'une bactérie aérobie dans un lac de très grande taille à la température de 297 K . Pour vivre, elle a besoin de consommer le dioxygène dissous dans l'eau au voisinage de sa surface. La bactérie est modélisée par une sphère de centre O fixe, de rayon R , sa masse volumique µ est assimilée à celle de l'eau. On se place en régime stationnaire et 3 on note n ( r ) la densité particulaire, exprimée en m , du dioxygène dissous à la distance r du centre O ( r > R ) . La diffusion du dioxygène dans l'eau obéit à la 9 2 1 loi de Fick avec un coefficient de diffusion D 2 10 m s . Loin de la bactérie, la concentration molaire volumique du dioxygène dissous dans le lac vaut 3 c 0 0, 2 mol m . On admet que la consommation en oxygène de la bactérie est proportionnelle à sa masse et on introduit le taux horaire A de consommation de dioxygène par 1 1 unité de masse, mesuré en mol kg s . I.A - Densité particulaire en dioxygène au voisinage de la bactérie I.A.1) Rappeler la loi de Fick reliant la densité de courant particulaire j = j ( r )e r à la densité particulaire n ( r ) . Concours Centrale-Supélec 2008 1/13 PHYSIQUE I Filière PC Filière PC I.A.2) Exprimer ( r ) le nombre de molécules de dioxygène entrant par unité de temps dans une sphère de rayon r ( r > R ) en fonction de j ( r ) et de r . Justifier que ne dépend pas, dans le cas étudié, du rayon r de la sphère considérée. I.A.3) Déterminer l'expression de la densité particulaire n 1 en dioxygène dis+ sous sur la surface extérieure de la bactérie ( n 1 = n ( R ) ) en fonction de , D , R , N A et de la concentration molaire volumique c0 de dioxygène à grande distance de la bactérie. I.B - Taille critique de la bactérie I.B.1) Exprimer en fonction de N A , A , de la masse volumique µ et du rayon R de la bactérie. I.B.2) En déduire l'expression de n 1 en fonction de µ , A , R , D , N A et c 0 . Commenter la variation de n 1 en fonction du rayon R de la bactérie. I.B.3) Quelle inégalité doit vérifier n 1 afin que la bactérie ne suffoque pas ? En déduire l'expression du rayon critique R c d'une bactérie aérobie en fonction de D , c 0 , µ et A . Vérifier l'homogénéité du résultat. Calculer numériquement 1 1 R c sachant que A = 0, 02mol kg s . Comparer ce résultat à la dimension caractéristique R = 1µm de la bactérie E. coli. I.B.4) Pour une bactérie sphérique de rayon critique R c , calculer la valeur numérique du nombre total de molécules de dioxygène consommées par unité de temps. Partie II - Propriétés électrostatiques de la membrane de la bactérie On considère une bactérie E.coli en suspension dans une solution aqueuse + (milieu extérieur) contenant des ions N a , Cl etc... La bactérie est limitée à sa périphérie par une couche continue de lipides et de protéines, appelée membrane, d'épaisseur d = 10nm . La membrane sépare le milieu interne de la bactérie, appelé cytoplasme, du milieu extérieur. Le cytoplasme est également une solution aqueuse contenant des ions, des molécules, des protéines, etc... Le cytoplasme et le milieu extérieur sont donc des électrolytes, c'est-à-dire des solutions conductrices contenant des cations et des anions. L'ensemble formé par le cytoplasme et le milieu extérieur est globalement neutre. Concours Centrale-Supélec 2008 2/13 PHYSIQUE I Filière PC II.A - Étude préliminaire : longueur de Debye dans un électrolyte Soit un électrolyte à la température T , globalement neutre, constitué d'une + solution aqueuse contenant majoritairement des ions N a et Cl avec un nom+ bre de cations N a par unité de volume uniforme n (densité volumique des cations). La permittivité de l'électrolyte est notée . On introduit dans cet électrolyte une plaque isolante chargée négativement ; les charges liées à la plaque sont réparties de manière uniforme sur sa surface. La plaque est confondue avec le plan x = 0 et l'électrolyte occupe le demi-espace x > 0 . Les effets de bord sont négligés. II.A.1) Après introduction de la plaque chargée, les densités volumiques des ions ne sont plus uniformes, pourquoi ? Représenter qualitativement sur un schéma la répartition des ions dans l'électrolyte. Que peut-on dire de la densité volumique de charge électrique dans une région proche de la plaque ? Dans une région très éloignée de la plaque ? + À présent, les densités volumiques des ions N a et Cl en un point M sont notées respectivement n + ( M ) et n ( M ) . Ces ions obéissent à la loi de Fick avec + des coefficients de diffusion D + pour N a et D pour Cl . II.A.2) Exprimer les vecteurs densités de courant volumique électrique j D+ et + j D associés respectivement à la diffusion des ions N a et Cl . II.A.3) Soumis à un champ électrostatique E , un ion acquiert une vitesse + limite µ ion E où le coefficient µ ion est sa mobilité. Les mobilités de N a et Cl sont notées µ + et µ - . Donner les expressions des vecteurs densités de courant volumique électrique j e+ et j e associés à l'action du champ électrostatique E + respectivement sur les ions N a et Cl . II.A.4) Écrire les équations de Maxwell de l'électrostatique dans l'électrolyte, en présence d'une densité volumique de charge due à un déséquilibre local entre le nombre de cations et le nombre d'anions ; on admettra que la prise en compte de la permittivité de l'électrolyte consiste à remplacer par la permittivité 0 du vide. Montrer que tout champ électrostatique E dérive d'un potentiel scalaire . Par la suite, on se place dans le cadre unidimensionnel, c'est-à-dire que tous les champs eulériens ne dépendent plus que de l'abscisse x (coordonnée cartésienne normale à la plaque chargée) : E = E ( x )e x ; = ( x ) ; n + = n + ( x ) ; n = n ( x ) . II.A.5) On suppose que l'équilibre est réalisé. a) Écrire pour chaque ion, une relation simple valable à l'équilibre, entre son vecteur densité de courant volumique électrique de diffusion et son vecteur densité de courant volumique électrique dû à l'action du champ électrostatique. En Concours Centrale-Supélec 2008 3/13 PHYSIQUE I Filière PC déduire, pour chaque ion, l'équation différentielle liant sa densité volumique au potentiel électrostatique ( x ) . b) Déterminer la loi de variation de la densité volumique de chaque ion en fonction de ( x ) . Le potentiel électrostatique sera supposé nul loin de la plaque : ( ) = lim ( x ) = 0 . x + c) En utilisant la relation d'Einstein qui relie la mobilité d'un ion à son coefficient de diffusion µ ± / D ± = ± e / k B T , où e est la charge élémentaire, k B est la constante de Boltzmann et T la température de l'électrolyte, montrer que les densités volumiques de ions peuvent finalement s'écrire : e ( x ) n + ( x ) = nexp --------------- kBT et e ( x ) n ( x ) = nexp + --------------- . kBT Quel nom donneriez-vous à ces lois de variation ? II.A.6) On désire déterminer la densité volumique de charge électrique ( x ) à l'équilibre. a) Exprimer ( x ) en fonction de ( x ) , n , e , k B et T . Simplifier cette expression dans l'hypothèse où e ( x ) « k B T . Dans la suite de cette partie, on utilisera l'expression simplifiée de ( x ) . b) Déduire de l'une des équations de Maxwell, une équation différentielle liant ( x) à ( x) . c) En déduire l'équation différentielle vérifiée par ( x ) . On fera apparaître une longueur caractéristique , appelée longueur de Debye, que l'on exprimera en fonction de e , n , (permittivité de l'électrolyte), k B et T . + d) Résoudre l'équation différentielle précédente sachant que E ( x = 0 ) = eff / où eff représente la densité surfacique de charge associée aux ions de l'électrolyte appartenant à l'interface solide/liquide en x = 0 . On exprimera la solution ( x ) en fonction de x , , et eff (que l'on ne cherchera pas à déterminer). Quel est le signe de eff ? e) En déduire la loi de variation de la densité volumique de charge ( x ) . Vérifier la conservation de la charge dans l'électrolyte. À quelle distance caractéristique de la plaque peut-on considérer que le potentiel électrostatique et la densité volumique de charge électrique sont négligeables ? Concours Centrale-Supélec 2008 4/13 PHYSIQUE I Filière PC II.B - Répartition des charges de part et d'autre de la membrane de la bactérie cytoplasme x1x 1 milieu extérieur membrane x2x 2 x3 x 3 x4x 4 x5x 5 x6x 6 xx E(x) E( x) 0 xx E0 ------E 0/2 2 -E E0 Figure 1 : Champ électrostatique La membrane de la bactérie permet des échanges d'ions et de molécules entre le cytoplasme et le milieu extérieur mais sa splitéabilité varie d'un ion à l'autre. À l'équilibre, la concentration de chaque ion est différente de part et d'autre de la membrane, il apparaît alors un champ électrostatique E . La figure 1 représente l'allure de la composante E ( x ) du champ électrostatique dans le modèle unidimensionnel ( E ( x ) 0 ) . Les domaines représentent : · le cytoplasme pour x x 2 · la membrane pour x 2 x x 5 · le milieu extérieur pour x x 5 Les permittivités du cytoplasme, de la membrane et du milieu extérieur sont supposées égales et ont la même valeur que la permittivité de l'eau. II.B.1) Tracer, sans effectuer de calcul mais en justifiant, l'allure du potentiel électrostatique ( x ) . II.B.2) Faire de même pour la distribution volumique de charge ( x ) . II.B.3) a) En utilisant les résultats de l'étude préliminaire II.A, donner l'expression de la dimension caractéristique d c des régions chargées. Concours Centrale-Supélec 2008 5/13 PHYSIQUE I Filière PC b) Au laboratoire, le milieu de culture utilisé pour étudier la bactérie 1 E. coli contient 5, 0 g L de NaCl (le milieu contient également d'autres ions + que N a et Cl mais avec des concentrations beaucoup plus faibles). Calculer numériquement d c et comparer le résultat à l'épaisseur d = 10nm de la membrane de la bactérie. Conclusion ? 1 Données : 80 0 (permittivité de l'eau), T = 297 K , M Na = 23, 0 g mol 1 (masse molaire de Na ) et M Cl = 35, 5g mol (masse molaire de Cl ). II.C - Potentiel de repos de la membrane de la bactérie La membrane de la bactérie possède une splitéabilité sélective suivant la + nature des ions. À l'intérieur du cytoplasme, les cations majoritaires sont K . Nous considérons à présent le modèle de Bernstein (1902) dans lequel seule la + diffusion des ions K est prise en compte, la membrane étant supposée imper+ méable à tous les autres ions. De plus, la densité volumique des ions K dans le cytoplasme est beaucoup plus élevée qu'à l'extérieur. On considère à nouveau le modèle unidimensionnel de la figure 1. II.C.1) Justifier l'existence d'un champ électrostatique E = E ( x )e x à l'équili+ bre, dû aux ions K . Dans quel sens est-il dirigé ? + II.C.2) La condition d'équilibre pour l'ion K conduit à l'équation différentielle suivante (résultat admis) : 1 dn e d ------------ ------- = ----------- -------n ( x ) dx k B T dx + où n ( x ) représente la densité volumique des ions K à l'abscisse x et ( x ) est le potentiel scalaire associé au champ électrostatique E = E ( x )e x . a) En déduire la différence de potentiel = int ext entre l'intérieur et l'extérieur de la bactérie, appelée potentiel de repos. Le résultat sera exprimé en fonction de k B , T , e , n int et n ext où n int et n ext représentent les concentrations + des ions K à l'intérieur et à l'extérieur de la bactérie. b) Sachant que n int / n ext = 100 pour la bactérie E. coli dans un milieu de culture à la température de 297 K , en déduire la valeur numérique de son potentiel de repos . Partie III - Propulsion d'un micro-organisme par flagelle Dans toute la suite du problème, on supposera que les nombres de Reynolds Re des écoulements sont petits devant 1 . L'observation d'une différence de direction entre la vitesse de sédimentation et la force de traînée s'exerçant sur un bâtonnet en mouvement dans un fluide visqueux a été à l'origine de la théorie de la propulsion des micro-organismes par le mouvement rotatif d'un flagelle. Concours Centrale-Supélec 2008 6/13 PHYSIQUE I Filière PC III.A - Chute d'un bâtonnet dans un fluide visqueux On considère un bâtonnet cylindrique à section circulaire ; ce bâtonnet présente donc une invariance par rotation autour de son axe que l'on notera Cz , C désignant le centre de masse du bâtonnet. La longueur L du bâtonnet est supposée grande devant son rayon. On notera P le poids apparent du bâtonnet, c'est-à-dire la somme de son poids et de la poussée d'Archimède exercée par le fluide. On réalise une série d'expériences de chutes du bâtonnet dans un fluide visqueux de viscosité . III.A.1) On observe qu'en régime permanent, la vitesse de chute v // , lorsque le bâtonnet est vertical, est égale au double de la vitesse de chute v lorsque le bâtonnet est horizontal. Dans les deux cas, la chute est un mouvement de translation verticale. Ceci conduit à une force de traînée qui s'exprime, pour un bâtonnet de longueur L , en fonction de la vitesse selon la loi : F x = v x ; F y = v y ; F z = // v z . Déterminer la valeur numérique du rapport = / // . III.A.2) Comment varient // et en fonction de la longueur L du bâtonnet ? On pourra supposer que // = K L , K étant une constante adimensionnée, et déterminer l'exposant par analyse dimensionnelle. III.A.3) En déduire que, pour un bâtonnet de longueur L , animé d'une vitesse v , dont la grande dimension est orientée selon le vecteur directeur u , la force de traînée peut s'écrire : F = KL ( 2v v // ) avec v // = ( v u )u . III.B - Propulsion d'une bactérie par flagelle ; aspect cinématique La propulsion de certaines bactéries est assurée par le mouvement de longs filaments appelés flagelles. Le flagelle de la bactérie E. coli est modélisé par un solide en forme d'hélice de rayon Figure 2 a = 0, 20 µm et de pas avec 2h Concours Centrale-Supélec 2008 7/13 PHYSIQUE I Filière PC h = 0, 10 µm . La longueur totale du filament est L = 10 µm . On note Oz l'axe du cylindre imaginaire sur lequel est enroulée cette hélice. Soit M ( ) le point courant de l'hélice ; on a, en coordonnées cylindriques : OM ( ) = ae r + he z . Un moteur biologique implanté dans la membrane de la bactérie peut imprimer 2 1 au flagelle un mouvement de rotation à la vitesse angulaire = 7, 0 10 rad s . 3 La bactérie est plongée dans l'eau de viscosité = 1, 0 10 Pa.s contenue dans un récipient immobile dans le référentiel L du laboratoire. La masse volumique de la bactérie est pratiquement égale à celle de l'eau. III.B.1) a) En supposant que le flagelle se visse dans le fluide environnant, de quelle distance d se propulse la bactérie pendant que le flagelle effectue un tour complet ? En déduire une première estimation de la vitesse de la bactérie. b) Vérifier la pertinence de l'hypothèse Re « 1 . III.B.2) La bactérie étant maintenue immobile dans le référentiel L du laboratoire, le flagelle est mis en rotation à la vitesse angulaire ( / L ) = e z dans le référentiel du laboratoire. a) Donner en coordonnées cylindriques l'expression dans L de la vitesse du point courant M du flagelle. b) Un tronçon du solide compris entre et + d est assimilé à un petit 2 2 bâtonnet de longueur dL = a + h d . Le vecteur unitaire u de l'axe de ce petit bâtonnet est : ( ae + he z ) u = ---------------------------2 2 a +h Préciser l'expression de la composante v // de la vitesse parallèle à l'axe du bâtonnet. III.C - Existence d'une force propulsive Les résultats du premier paragraphe, avec K = 4 , conduisent pour un tronçon de flagelle de longueur dL à une force de traînée dF = 4 ( 2v v // )dL . III.C.1) La bactérie est maintenue immobile, et le flagelle est en rotation à la vitesse angulaire ( / L ) = e z dans le référentiel L du laboratoire. a) Exprimer la force de traînée dF 1 s'exerçant sur ce bâtonnet élémentaire. b) En supposant que varie de 2 p à 0 où p = L / ( 2a ) est entier, montrer que la force de traînée exercée par le fluide environnant sur le flagelle entier est Lah F 1 = 4 ---------------------- e z .En déduire que, pour un signe approprié de , la 2 2 a +h Concours Centrale-Supélec 2008 8/13 PHYSIQUE I Filière PC bactérie peut se propulser selon e z . III.C.2) Lorsque le flagelle est en mouvement de translation à la vitesse ve z par rapport au référentiel L on montre, par un raisonnement analogue au précédent, que la force de traînée F 2 exercée par le fluide environnant sur le flagelle entier est 2 2 h + 2a F 2 = 4 Lv -------------------------- e z . 2 2 a a +h (résultat admis) En remarquant la linéarité de la relation entre la force de traînée et la vitesse, quelle est la force de traînée F exercée par le fluide environnant sur le flagelle entier lorsqu'il tourne à la vitesse angulaire e z tandis que la bactérie se propulse à la vitesse ve z ? III.C.3) La bactérie (hors flagelle) est modélisée par une sphère de rayon R = 0, 5µm . On rappelle l'expression de la traînée s'exerçant sur une sphère de rayon R en mouvement à la vitesse v dans un fluide de viscosité : T = 6Rv . a) Montrer que cette force peut être négligée devant F 2 . b) Justifier l'estimation de la vitesse de la bactérie en régime permanent : 2 a v p = h --------------------- e et calculer numériquement cette vitesse. 2 2 z 2a + h III.D - Aspect énergétique III.D.1) Exprimer en fonction des forces dF 1 et dF 2 définies au paragraphe précédent le moment élémentaire d M O en O des efforts exercés par le fluide sur un tronçon [ , + d ] du flagelle. III.D.2) Lorsque la bactérie est propulsée par le flagelle à la vitesse v p , on obtient par sommation une composante sur e z de ce moment 2 Mz 2 3/2 aL ( a + h ) = 8 -------------------------------------- . 2 2 2a + h (résultat admis) Calculer M z numériquement. III.D.3) Déterminer littéralement puis numériquement la puissance P m du moteur cellulaire qui maintient le flagelle en rotation à la vitesse angulaire 2 1 = 7, 0 10 rad s . Concours Centrale-Supélec 2008 9/13 PHYSIQUE I Filière PC Partie IV - Moteur rotatif biologique La rotation du flagelle est assurée par un moteur rotatif biologique de taille nanométrique, incorporé dans la membrane de la bactérie. L'objectif de cette partie est d'étudier quelques caractéristiques de ce moteur rotatif. IV.A - Source d'énergie du moteur IV.A.1) Question préliminaire Soit G = U + pV TS l'enthalpie libre d'un système thermodynamique d'énergie interne U , de volume V , d'entropie S à la température T et sous la pression p . On rappelle l'inégalité vérifiée par le travail utile récupérable W (autre que celui des forces de pression) et la variation d'enthalpie libre G du système entre un état d'équilibre initial et un état d'équilibre final au cours d'une évolution monotherme et monobare : W G . À quelle condition l'inégalité précédente devient-elle une égalité ? On considère le système fermé constitué de la bactérie et du milieu aqueux (milieu extérieur) dans lequel elle évolue. La température T et la pression p du système, grandeurs uniformes, sont maintenues constantes. On admet que la différentielle de l'enthalpie libre du système s'écrit dans cette situation dG = ( µ e + e e )dn e + ( µ i + e i )dn i , où : · dn e et dn i représentent respectivement les variations du nombre de protons + H à l'extérieur et dans le cytoplasme (milieu intérieur) de la bactérie ; · e et i sont respectivement les potentiels électrostatiques du milieu extérieur et du cytoplasme ( e est la charge élémentaire) ; + [H ] c° + [H ] c° · µ e = µ° + k B T ln --------------e- et µ i = µ° + k B T ln --------------i , où : · le terme µ° ne dépendant que de T est une constante ; · k B est la constante de Boltzmann ; + + · [ H ] e et [ H ] i sont les concentrations molaires des protons à l'extérieur et dans le cytoplasme ; 1 · c° = 1, 0 mol L est une concentration de référence. IV.A.2) Les protons en traversant la membrane fournissent l'énergie nécessaire au fonctionnement du moteur. Montrer que le travail utile récupérable W lorsque dn protons traversent la membrane dans le sens milieu extérieur cytoplasme s'écrit, à la limite du fonctionnement réversible : W = epdn où p , grandeur appelée force protomotrice, sera exprimée en fonction de e , k B , T , + + [ H ] e , [ H ] i et = i e . Concours Centrale-Supélec 2008 10/13 PHYSIQUE I Filière PC IV.A.3) En déduire qu'à la température de 297 K , la force protomotrice peut également s'écrire : p = 0, 059pH + ( V ) où pH = pH i pH e avec + + pH i = log ( [ H ] i / c° ) et pH e = log ( [ H ] e / c° ) . IV.A.4) Calculer p à 297 K sachant que la bactérie E. coli maintient pH i = 7, 8 à l'intérieur de son cytoplasme, que le milieu extérieur impose pH e = 7, 0 et que = 0, 12 V . IV.B - Caractéristique couple-fréquence de rotation On désire étudier la loi de flagelle raccourci variation du couple délivré bille de latex par le moteur rotatif en fonction de la fréquence de rota tion f du flagelle. À cette fin, on considère une bactérie bactérie E. coli, ayant la forme d'un bâtonnet cylindrique aux surface en verre Figure 3 extrémités arrondies, liée à la surface en verre de l'une des faces du récipient qui la contient. En outre, la longueur du flagelle a été considérablement réduite par un procédé chimique et on y a attaché une petite bille de latex de rayon micrométrique a 0 (cf. figure 3). Lorsque la petite bille de latex tourne à la vitesse de rotation uniforme = 2f autour d'un axe passant par son centre, elle subit un couple de frottement fluide 3 v = 8a 0 résultant de l'action des forces de viscosité où est la viscosité dynamique de la solution dans laquelle plonge la bactérie. Le couple visqueux s'exerçant sur le flagelle raccourci sera négligé. IV.B.1) Montrer que v est bien homogène à un couple. Donner l'expression, en régime permanent, du couple moteur en fonction de a 0 , et . IV.B.2) La fréquence de rotation f de la bille de latex peut être mesurée par un procédé d'optique physique. Sur quel(s) paramètre(s) peut-on jouer pour obtenir le tracé de la caractéristique en fonction de f ? L'allure de la caractéristique expérimentale ( f ) est fournie sur la figure 4. On y distingue deux régimes de fonctionnement : · un bas régime ( 0 f 175 Hz ) où le couple moteur est pratiquement constant ; · un haut régime ( 175 f 350 Hz ) où le couple moteur chute avec la fréquence de rotation. Concours Centrale-Supélec 2008 11/13 PHYSIQUE I Filière PC / / max max Figure 4 1 bas régime haut régime 0 0 50 100 150 175 200 250 300 350 f (Hz) IV.B.3) Déterminer la valeur numérique du couple maximal max , sachant qu'une petite bille de latex de rayon a 0 = 0, 50µm tourne à la fréquence f = 10 Hz dans une solution de viscosité = 0, 020 Pa s . IV.B.4) Donner l'ordre de grandeur numérique de la fréquence de rotation du flagelle raccourci lorsqu'aucune bille de latex n'est accrochée. IV.C - Évaluation du rendement du moteur On considère toujours la même bille de latex entraînée à la fréquence de rotation f = 10 Hz par le moteur de la bactérie dans une solution de viscosité = 0, 020 Pa s . IV.C.1) Déterminer l'expression de la puissance mécanique P m fournie par le moteur. Faire l'application numérique avec les données précédentes. IV.C.2) Sachant que pour chaque tour effectué par le moteur, le nombre de protons traversant la membrane de la bactérie est n = 1200 , déterminer l'expression littérale du flux de protons traversant la membrane en fonction de n et f . Faire l'application numérique. IV.C.3) En utilisant le résultat de la question IV.A.2, exprimer la puissance Pr reçue par le moteur due à la traversée de la membrane par les protons en fonction de la force protomotrice p , de la charge élémentaire e et du flux des protons . Faire l'application numérique sachant que p = 0, 17 V . IV.C.4) Définir et calculer la valeur du rendement de ce moteur rotatif biologique pour la fréquence f = 10 Hz . Comment évolue le rendement lorsque la fréquence de rotation f varie dans le domaine de fonctionnement bas régime ? Dans le domaine de fonctionnement haut régime ? Concours Centrale-Supélec 2008 12/13 PHYSIQUE I Filière PC IV.D - « Moteur pas à pas » Le rotor du moteur U.B. est assimilé à un F F disque de rayon U.B. U.B. r = 20 nm et rotor F F axe de rotation : d'épaisseur négli geable. La rotation U.B. « ``unité biologique''»: U.B. du rotor est assu- U.B. r rée par un stator F F constitué de huit « unités U.B. U.B. biologiques », F U.B. F Figure 5 régulièrement réparties sur sa périphérie et générant chacune une force orthoradiale d'intensité F (cf. figure 5). On envisage le fonctionnement du moteur en bas régime. IV.D.1) Exprimer l'intensité F de la force en fonction de r et max . Calculer numériquement F . Afin d'avoir une idée intuitive de ce que représente cette force, calculer numériquement la distance qui doit séparer deux électrons dans le vide pour obtenir une force électrostatique de même intensité. IV.D.2) Le moteur biologique est en fait un « moteur pas à pas », c'est-à-dire qu'il tourne par bonds successifs d'un angle fixe dit incrément. Dans notre cas, un tour complet s'effectue en 52 incréments. Calculer le déplacement d d'un point de la périphérie du rotor correspondant à un incrément. En déduire la valeur numérique du travail fourni par une « unité biologique » pour réaliser un incrément. IV.D.3) Déterminer le nombre de protons n qui doivent traverser la membrane pour qu'une « unité biologique » réalise un incrément. On supposera que le rendement du moteur est de 100% pour ce calcul. ··· FIN ··· Concours Centrale-Supélec 2008 13/13